BOLOR VERDE DA
LARANJA (Citrus sinensis) CAUSADO POR Penicillium
digitatum
Érica de
Castro Costa
Acadêmico do curso de Agronomia
INTRODUÇÃO
A laranja (Citrus sinensis L. Osbeck – Rutaceae) fruto da laranjeira é uma deliciosa fruta cítrica e
uma das frutas mais cultivadas no mundo todo. Sua
origem é controversa, diz-se que pode ser oriunda da Índia, China ou Vietnã,
mas sabe-se de fato que a laranja é consumida desde à antiguidade. Seu tronco e
ramos apresentam casca castanho-acinzentada e são um tanto tortuosos. As folhas
são verdes, coriáceas, brilhantes e muito aromáticas (PATRO, 2013).
A
frutificação pode-se estender durante todo o ano, mas é mais abundante no
outono. Há cerca de 100 variedades de laranja produzidas em escala comercial.
No Brasil, as diferentes variedades de laranja são classificadas em três grupos
principais. As laranjas de umbigo, próprias para a mesa, caracterizadas pela
presença de um fruto rudimentar no ápice do fruto; as laranjas comuns, mas
ácidas e próprias para sucos; e as laranjas de baixa acidez, como a
laranja-do-céu, de sabor mais suave, muito doce e pouco ácida (PATRO, 2013).
O Brasil é o maior produtor mundial de laranja, respondendo
por cerca de 30 % da produção da fruta, 50% da produção de suco, com
participação de 85 % do suco comercializado internacionalmente. O cultivo de
espécies cítricas está assentado em uma área aproximada de 839 mil hectares,
dos quais 77 % se encontram localizados na região Sudeste do País. Cerca de 80 %
da produção de laranja no Estado de São Paulo é destinada à industrialização,
cujo suco produzido é exportado para países como Rússia, Bélgica, Países
Baixos, Estados Unidos e Japão, sendo os 20 % restantes destinados ao consumo “in natura”, tanto para mercado interno como para exportação
segundo o AGRIANUAL de 2011 (KUPPER et al., 2013).
A exportação
brasileira de laranja in natura não
atinge 0,5 % da produção nacional, sendo, portanto, marginal. Mesmo assim,
lidera em volume, tendo alcançado 99,2 mil toneladas, que renderam ao país 20,4
milhões de dólares em 1996. Problemas com variedades, doenças, etapas
pós-colheita e embalagens contribuem para resultados pouco expressivos (FRANCO e
BETTIOL, 2000).
Doenças
em pós-colheita reduzem a quantidade e a qualidade de frutos comercializáveis
e, em citros, podem provocar grandes perdas (FORNER et al., 2013). O bolor-verde é a principal doença pós-colheita dos
citros e está disseminada em todos os países produtores, afetando todas as
espécies cítricas. Sua ocorrência depende muito das condições climáticas e da
forma de manipulação dos frutos, desde o pomar até a chegada ao consumidor,
podendo ocorrer tanto no armazenamento como no transporte do produto (LARANJEIRA
et al., 2005).
O bolor-verde é
uma podridão mole dos frutos com posterior cobertura da lesão pelo micélio
branco e por grande número de esporos verdes do fungo. Os esporos maduros
soltam-se com grande facilidade a qualquer movimento ou impacto sobre os frutos
afetados. Assim, são facilmente dispersos pelo manuseio, pela agitação ou
exposição dos frutos à correntes de ar. A infecção é dependente de ferimentos,
podendo ocorrer em qualquer etapa a partir da maturação dos frutos.
Temperaturas em torno de 24 ºC são consideradas ótimas e maiores que 30 ºC ou
menores que 5-7 ºC paralisam o crescimento do fungo. Em “packing houses”, devido à presença constante de frutos
sadios e doentes, os ciclos de infecção e esporulação podem ocorrer repetidas
vezes no período de colheita (LARANJEIRA
et al., 2005).
O gênero Penicillium possui
três espécies de importância na podridão de frutos cítricos, a saber: P. digitatum, P. italicum e P. ulaiense. Esse gênero é caracterizado
por possuir conidióforos que surgem do micélio ou únicos (P. digitatum e P. italicum)
ou em forma de sinêmios (P. ulaiense),
ramificados próximos ao ápice, penicilados (formato de vassoura), terminando em
um grupo de fiálides. Conídios (fialósporos) hialinos ou ligeiramente
coloridos, quando agrupados, formam massas, são unicelulares e, na sua maioria,
globosos ou ovoides, formando no seco, cadeias basipetais (LARANJEIRA et al., 2004).
De acordo com a
pesquisa realizada no Banco de Dados - Fungus Host Distribution Database (2015)
tem sido registrado a ocorrência do patógeno em 17 gêneros e 50 espécies de
plantas hospedeiras representadas por P.
digitatum mundialmente. Infectando
a espécie de laranja (C. sinensis),
nove espécies foram identificadas e
representadas por P. citrinum, P.
digitatum, P. fructigenum, P. glaucum, P.
italicum, P. lutem, P. roseum, P. ulaiense e P. verrucosum (FARR & ROSSMAN,
2015).
Como forma de
controlar esse patógeno vêm sendo utilizados o tratamento químico, a
irradiação, a termoterapia, filmes plásticos e práticas culturais para reduzir
o inóculo no campo. Os fungicidas do grupo dos benzimidazóis são os mais
utilizados no Brasil. Entretanto, possuem várias restrições de uso, como a de
selecionar estirpes de Penicillium sp.
resistentes ao fungicida. Dessa forma, novos fungicidas vêm sendo estudados,
mas ainda não possuem registro para uso em pós-colheita no Brasil. A situação
se agrava no caso de frutos para exportação, uma vez que existem diferenças
entre as legislações dos países consumidores na aceitação de determinados
fungicidas, bem como na concentração de resíduos tolerada (FRANCO e BETTIOL,
2000).
Objetivo
O objetivo deste boletim técnico é descrever a
sintomatologia, etiologia, epidemiologia e controle do bolor-verde da laranja
causado por Penicillium digitatum.
DESENVOLVIMENTO
Hospedeiro/cultura:
Laranja (Citrus sinensis L. Osbeck)
Família
Botânica: Rutaceae
Doença:
Bolor-verde
Agente
Causal (Teleomorfo): Penicillium digitatum Sacc. (1881)
Local
de Coleta: Orizona, GO
Data
de Coleta: 17/03/2015
Taxonomia:
A fase
teleomórfica pertence ao reino Fungi, filo Ascomycota, classe Eurotiomycetes,
ordem Eurotiales, família Trichocomaceae. A fase anamórfica pertence ao Reino
Fungi, grupo incerto dos Fungos Mitospóricos, subgrupo dos hifomicetos, gênero Penicillium sp., espécie Penicillium digitatum (INDEX FUNGORUM,
2015).
Sintomatologia:
O
fruto analisado estava com cerca de 50 % de área total infectada e apresentava
seis cm de diâmetro de lesão, apresentando parte de micélio de coloração
verde-oliva (Fig. 1A) representada pela produção de conídios, sendo circundada
por uma de coloração esbranquiçada representada pelo micélio vegetativo (Fig.
1A). Em menos de sete dias, toda a área do tecido encontrava-se colonizada pelo
patógeno, notando-se odor característico de azedo.
Os
bolores causam podridões moles em frutos (Fig. 1A), que se iniciam por pequenas
anasarcas na superfície da casca e que, rapidamente, aumentam de tamanho até
tomarem todo o fruto. O fungo desenvolve um micélio branco (Fig. 1A) sobre o
tecido afetado, que depois é revestido por uma densa massa de esporos, cuja cor
varia em função do fungo envolvido. No bolor verde o revestimento é de cor
verde-oliva e no bolor azul ele é azul no início, podendo depois adquirir a cor
marrom-oliva. No bolor verde, o desenvolvimento das lesões é mais rápido. As
massas de esporos verdes (Fig. 1A) são circundadas por uma ampla faixa de
crescimento fúngico branco, que é separada da área sadia do fruto por uma
estreita camada de tecido de casca encharcado ao seu redor é mais pronunciada.
É comum encontrar frutos que apresentam mais de um tipo de bolor. Em condições
de elevada umidade, as podridões moles provocam uma rápida e completa
desintegração no fruto. Em ambiente seco, os frutos infectados murcham e
mumificam (FEICHTENBERGER et al.,
2005).
O
primeiro estádio manifesta-se pelo surgimento de uma mancha circular mole de
aspecto encharcado e pela ligeira descoloração da superfície afetada (Fig. 1A).
Esta área mede aproximadamente 0,5 centímetros e cede facilmente à pressão dos
dedos. O segundo estádio compreende o aparecimento de micélio na superfície do
fruto e o desenvolvimento, no centro do micélio, de uma massa de esporos
verdes. Em temperaturas favoráveis, em menos de dois dias, a lesão alcança
tamanho entre quatro a cinco centímetros de diâmetro, atingindo as vesículas de
suco. Circundando a esporulação verde (Fig. 1A) do patógeno, há uma faixa de
micélio branco, e externamente a ela, uma região enrugada na casca, de
consistência macia. A cor dos esporos é visível (Fig. 1A) geralmente em lesões
de diâmetro maior que dois centímetros. Eles são inicialmente verde-claros,
passando a verde-oliva à medida que envelhecem (LARANJEIRA et al., 2005).
Etiologia
(Sinais):
As colônias (Fig. 1C) desse fungo desenvolvidas em
meio Czapek tem uma aparência similar, apresentando um crescimento muito
restrito, porém rápido. Possui superfície plana e pode formar uma cultura de
coloração verde-alaranjado a cinza-oliváceo; forma um odor bastante forte; e
desencadeia a queda do fruto. No verso da placa, tem-se uma coloração de palha
a amarronzada ou oliva (ONIONS, 1966). Para Samson et al. (2010), as colônias tem crescimento lento ou muito rápido em
meio CYA à 25 ºC (diâmetro de 25-55 mm depois de sete dias), apresentando
coloração amarelo ao marrom-esverdeado e superfície aveludada. No verso da
placa apresenta colônia não sulcada e de coloração pálida a bege. Em meio MEA o
conídio apresenta coloração amarela a castanho-esverdeado, com colônias
velutinas, e no verso da placa apresenta uma coloração pálida. Em meio YES-agar
o grau de esporulação é forte, de coloração bege para avelã no verso da placa,
e apresentam crescimento rápido (diâmetro de 44-76 mm depois de sete dias). Em
meio CREA, o crescimento é muito pobre e não há produção de ácido.
Os conidióforos (Fig. 1E) surgem da espécie ou hifas
aéreas, estipes de 70 a 150 µm de comprimento, com paredes finas, lisas e
terminando em penicílio; quando bem desenvolvidas são triverticilados, embora
mais frequente biverticilados ou irregulares (LARANJEIRA et al., 2004). São pequenos e apresentam dimensões de 30 – 100 x
4-5 µm, são lisos e apresentam uma constrição na base ou um micélio sub
emergente (ONIONS, 1966). A métula é variável possuindo 15-30 x 4-6 µm. As fiálides
representam um tipo de célula conidiogênica. Quando está bem desenvolvida ela
assemelha-se a uma vassoura (Fig. 1F). Podem possuir um formato assimétrico,
irregular, apresentando grande variação do número e das dimensões das suas
partes, e com poucas ramificações. Apresentam dimensões de 15-28 x 3-5 µm. Produzem
conídios em cadeia (Fig. 1H), de formato elíptico. (ONIONS, 1966).
Os conídios são variáveis, lisos, subglobosos
ou cilíndricos, mas usualmente, apresentam formato elíptico (Fig. 1I). Possuem dimensões
de 3,4-12 x 3-8 µm, usualmente apresentam 6-8 x 4-6 µm (ONIONS, 1966). Já para Laranjeira et al. (2005) o tamanho dos conídios
varia de 4 a 7 µm x 6 a 8 µm, com formato ligeiramente cilíndrico a ovalados
(Fig. 1I), sendo catenulados (Fig. 1H) variando ligeiramente na forma e no
tamanho, mesmo sendo originários do mesmo cordão de esporos. São muito frágeis
e desprende das células conidiogênicas com facilidade.
Samson et al. (2010) descreve a micro
morfometria de P. digitatum com conidióforos
hialinos, irregulares, ramificados, consistindo de pequenas estirpes, 60-160 x
5,0-7,0 µm, com poucas métulas e ramos terminados em verticilos de 3-6
fiálides, cilíndrica e dimensões de 15-25 x 5-6 µm. As fiálides são muitas
vezes solitárias, cilíndricas e curtas, de tamanho variável 10-20 x 4,0-5,0 µm.
Os conídios são elipsoides a cilíndricos, de parede lisa, apresentam coloração
verde-oliva quando em massa, de tamanho variável, mas principalmente 3,5-8 x
3,0-4,0 µm.
Tabela
1. Aspectos
comparativos da morfologia e morfometria do isolado de P. digitatum com os isolados descritos por Onions (1966) e Samson et al. (2010).
|
Características taxonômicas
|
Isolado Urutaí, GO (2015)
|
P. digitatum (Onions, 1966)
|
P. digitatum (Samson et al., 2010)
|
|
|
|
Coloração da colônia:
|
verde oliva a cinza oliváceo, com bordas esbranquiçadas (meio
BDA)
|
verde alaranjado a cinza-oliváceo
|
CYA - amarelo ao marrom-esverdeado; MEA - amarelo a
castanho-esverdeado; YES agar - bege para avelã
|
|
|
Características do Conidióforo:
|
|
|
|
|
|
Dimensão (µm):
|
nd
|
30-100 x 4-5
|
60-160 x 5,0-7,0
|
|
|
Coloração:
|
Hialina
|
nd
|
Hialina
|
|
|
Ramificação:
|
poucas, geralmente 3 ramos
|
pouca ramificação
|
poucas métulas e ramos
|
|
|
Métula:
|
Presente
|
presente
|
Presente
|
|
|
Dimensão (µm):
|
nd
|
15-30 x 4-6
|
15-25 x 5-6
|
|
|
Fiálide:
|
Presente
|
presente
|
presente (3 a 6)
|
|
|
Dimensão (µm):
|
Nd
|
15-28 x 3-5
|
10-20 x 4-5
|
|
|
Formato:
|
assimétrico, irregular
|
assimétrico, irregular
|
Cilíndricas
|
|
|
Características dos Conídios:
|
|
|
|
|
|
Dimensão (µm):
|
3,43-(6,99)-8,89 x 2,54-(3,93)-6,78
|
6-8 x 4-6
|
3,5-8 x 3-4
|
|
|
Formato:
|
ovóides e elípticos
|
subglobosos, cilíndricos ou elípticos
|
elipsoides e cilíndricos
|
|
|
Produção de conídios:
|
catenulados
|
catenulados
|
Nd
|
|

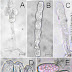
Figura 1. Sintomas e sinais do
bolor verde da laranja (Citrus sinensis) causado por Penicillium digitatum. A. sintoma da doença na superfície de
um fruto maduro (bar = 13,1 mm), B. fruto
contaminado após seis dias do teste de inoculação (bar = 23,9 mm), C. cultura em meio BDA nos quatro dias
após a inoculação (bar = 23,6 mm), D. detalhe
da margem esbranquiçada da cultura (bar = 3,5 mm), E. partes do conidióforo (bar = 8,14 µm), e1. estipe (ramo), e2. métula,
F. (bar = 4,57 µm), G. célula conidiogênica fialídica
(seta)(bar = 14,91 µm), H. conídios
em cadeia basipetal (bar = 3,48), I. conídios
ovóides e esféricos (seta)(bar = 12,22 µm).
Epidemiologia:
O bolor
esverdeado dos frutos de laranja cresce rapidamente. A superfície dos frutos se
tornam cobertos por uma massa branca inicialmente, e depois se tornam um
verde-oliva a coloração devido a produção de conídios. O P. digitatum se diferencia de P.
italicum, que apresenta uma coloração verde azulada e que pode reduzir a
massa do fruto. O bolor verde é comum em todas as áreas de plantio de citros e
é espalhado principalmente em frutos, causando a podridão dos mesmos em
condições de armazenamento (ONIONS, 1966).
Eles sobrevivem saprofiticamente, em pomares e outros ambientes, sobre
vários tipos de substratos orgânicos, na forma de conídios (FEICHTENBERGER et al., 2005). A infecção depende de
ferimentos, podendo ocorrer em qualquer etapa a partir da maturação dos frutos.
Além dos ferimentos, a temperatura é o outro fator importante para o desenvolvimento
do bolor-verde. Temperaturas em torno de 24ºC são consideradas ótimas e maiores
que 30 ºC ou menores que 5-7 ºC paralisam o crescimento do fungo. Em geral, à
medida que ocorre diminuição da temperatura, o desenvolvimento do fungo é mais
lento, e, portanto, os sintomas são menos evidentes (LARANJEIRA et al., 2005).
Em
“packing houses”, devido à presença constante de frutos sadios e doentes, os
ciclos de infecção e esporulação podem ocorrer repetidas vezes no período de
colheita. Em contêineres, a infecção não ocorre de um fruto doente para um
sadio adjacente, já que não há transmissão por contato (LARANJEIRA et al., 2005).
O bolor verde da
laranja possui como agente causal a espécie P.
digitatum descrita por Sacc., e publicada em Fungi italica autogr.
del. 17-28: tab. 894 no ano de 1881. Apresenta 205 registros de
patogenicidade infectando 17 gêneros e 50 espécies de plantas hospedeiras,
distribuídas mundialmente (FARR & ROSSMAN, 2015).
Destas,
34 espécies pertencem ao gênero Citrus, são
elas: Citrus aurantifolia registrado
nos EUA (French, A.M. 1987) (Alfieri Jr. et. al., 1984), em Cuba (Arnold, G.R.W. 1986), na Venezuela (Urtiaga, R. 2004), C. aurantifolia var mexicana registrado
em Cuba (Urtiaga, R. 1986), na Venezuela (Urtiaga, R. 1986), C.
aurantium registrado em Barbados (Norse, D. 1974), nos EUA (Alfieri Jr., S.A. et. al., 1984), na Grécia
(Pantidou, M.E. 1973), em Myanmar (Thaung, M.M. 2008), na África do Sul (Gorter, G.J.M.A. 1977), na Espanha (Gonzalez Fragoso, R. 1916). C. chrysocarpa foi registrado na Índia (Rao, V.G. 1969).
C.
deliciosa, registrado
na Grécia (Pantidou, M.E. 1973),
C.
grandis registrado
nos EUA (French, A.M. 1987),
na China (Tai, F.L. 1979),
no Japão (Kobayashi, T. 2007);
C.
iyo registrado
no Japão (Kobayashi, T. 2007);
C.
jambhiri registrado
nos EUA (Alfieri Jr., S.A. et. al.,
1984); C.
junos registrado
na China (Tai, F.L. 1979),
no Japão (Kobayashi, T. 2007);
C.
latifolia registrado
nos EUA (French, A.M. 1987); C. limon registrado na Austrália (Shivas, R.G. 1989), nos EUA (French, A.M. 1987)(Alfieri
Jr. et. al., 1984); na China (Tai,
F.L. 1979), em Cuba (Urtiaga,
R. 1986), em Greece
(Pantidou, M.E. 1973), em Hong Kong (Lu, B. et. al., 2000), na Itália (Balmas, V. et. al., 2005), no Japão
(Kobayashi, T. 2007), no Malawi (Peregrine, W.T.H., and Siddiqi, M.A. 1972), no México (Alvarez, M.G. 1976), Nova
Zelândia (Pennycook, S.R. 1989),
no Niue (Dingley, J.M. et. al., 1981),
na Tanzânia (Riley, E.A. 1960)
e no Zimbábue (Riley, E.A. 1960).
C.
limonia registrado no Chile (Mujica, F., and Vergara, C. 1945), na China (Tai, F.L. 1979), nos EUA (Alfieri Jr., S.A. et. al., 1984),
Serra Leoa (Deighton, F.C. 1936),
África do sul (Gorter, G.J.M.A. 1977),
África do Sul (Doidge, E.M. 1950).
C.
limonium registrado
em Cyprus (Georghiou, G.P., and
Papadopoulos, C. 1957), na República Dominicana (Ciferri, R. 1961). C. maxima registrado em Barbados (Norse, D. 1974), nos EUA(French, A.M. 1989) (Alfieri
Jr., S.A. et. al., 1984), na África do Sul (Doidge, E.M. 1950). C. medica registrado em Brunei Darussalam (Peregrine, W.T.H., and Ahmad, K.B. 1982), na China (Tai, F.L. 1979), nos EUA (Alfieri Jr., S.A. et. al., 1984) e na Grécia
(Pantidou, M.E. 1973). C. meyerii registrado nos EUA (French,
A.M. 1987); C. microcarpa registrado na China (Tai,
F.L. 1979); C. natsudaidai registrado no Japão (Kobayashi,
T. 2007); C. nobilis registrado em Brunei Darussalam (Peregrine, W.T.H., and Ahmad, K.B. 1982), na China (Tai, F.L. 1979) e no Japão (Kobayashi, T. 2007). Citrus
nobilis var. deliciosa
registrado
na Serra Leoa (Deighton,
F.C. 1936) na África do Sul (Doidge, E.M. 1950). C. reticulata registrado na Austrália (Simmonds, J.H. 1966), nos EUA (French, A.M. 1987) (Alfieri
Jr., S.A. et. al., 1984), China (Tai,
F.L. 1979), em Cuba (Arnold,
G.R.W. 1986), em Hong Kong
(Lu, B. et. al., 2000), na Índia (Gurunath Rao, V. 1966), no Japão
(Kobayashi, T. 2007), na
Nova Zelândia (Pennycook, S.R. 1989),
na África do Sul (Gorter, G.J.M.A. 1977),
na Venezuela (Urtiaga, R. 2004).
C.
reticulata var. kinokuni registrado na China (Tai, F.L. 1979); C.
reticulata var. poonensis registrado
na China (Tai, F.L. 1979), C.
reticulata var. suavissima registrado
na China (Tai, F.L. 1979), C.
reticulata var. suhoiensis registrado na China (Tai, F.L. 1979),
C.
reticulata var. tankan registrado
na China (Tai, F.L. 1979), C. sinensis registrado na Austrália (Shivas, R.G. 1989), no Brasil (Mendes, M.A.S. et. al., 1998), nos EUA (French, A.M. 1987),
no Chile (Mujica, F., and Oehrens, B.E. 1967),
na China (Tai, F.L. 1979),
em Cook Islands (Dingley, J.M et. al.,
1981), em Cuba (Arnold, G.R.W. 1986), na República Dominicana (Ciferri, R. 1961), no Fiji (Firman, I.D. 1972), nos EUA (Alfieri Jr., S.A. et. al., 1984), na Itália
(Licciardello, G. et. al., 2006), no Japão (Kobayashi, T. 2007), na
Malásia (Johnston, A. 1960),
no México (Alvarez, M.G. 19
76), na Nova Zelândia (Pennycook, S.R. 1989),
no Porto Rico (Stevenson, J.A. 1975),
na África do Sul (Gorter, G.J.M.A. 1977),
na Venezuela (Urtiaga, R. 1986)
e na Zambia (Riley, E.A. 1956).
C.
sinensis
var. brasiliensis registrado
no Japão (Kobayashi, T. 2007),
Citrus sp. registrado na Austrália (Cunnington, J. 2003), no Brasil (Mendes, M.A.S et.al., 1998), na Califórnia (El-Goorani, M.A., and
Sommer, N.F. 1979), na Costa
Rica (McGuire Jr.,
J.U., and Crandall, B.S. 1967), El Salvador
(McGuire Jr., J.U., and Crandall, B.S. 1967),
no Japão (Kobayashi, T. 2007),
no México (McGuire Jr., J.U., and
Crandall, B.S. 1967), na Nova Zelândia (Pennycook, S.R. 1989), no Panamá (McGuire Jr., J.U., and Crandall, B.S. 1967), Nova Guiné (Shaw, D.E. 1984) e no Porto Rico
(Stevenson, J.A. 1975.). C. tangerina registrado na China (Tai, F.L. 1979), C. tankan registrado na China (Zhuang,
W.-Y., Ed. 2001), C. unshiu registrado no Japão (Kobayashi,
T. 2007) e na Coreia (Cho,
W.D., and Shin, H.D., Eds. 2004). C.
×paradisi registrado
nos EUA (French, A.M. 1987)
(Alfieri Jr., S.A. et. al., 1984), Gana
(Dade, H.A. 1940), no Japão (Kobayashi, T. 2007), Porto Rico (Stevenson, J.A. 1975), na África do Sul (Gorter, G.J.M.A. 1977), na
Tanzânia (Riley, E.A. 1960),
Ilhas Virgens (Stevenson, J.A. 1975)
e no Zimbábue (Whiteside, J.O. 1966).
C.
×tangelo registrado
nos EUA (French, A.M. 1987)
e na Nova Zelândia (Pennycook, S.R. 1989)
(FARR & ROSSMAN, 2015).
Causando doenças
à hospedeira laranja (Citrus sinensis),
encontram-se registradas nove espécies de Penicillium
representados por P. fructigenum, P. glaucum,
P. italicum. P. lutem, P. roseum, P. ulaiense e P. verrucosum, e as que ocorrem no Brasil: P. citrinum e P. digitatum (FARR & ROSSMAN, 2015).
Foram encontrados
registros de doenças causadas por P. digitatum em seis hospedeiros diferentes no
Brasil, sendo eles o cajueiro, melão, feijão-fradinho, e diversas variedades de
citros. Há registro do patógeno causando doenças chamadas de bolor verde, mofo
verde, podridão verde do fruto e podridão do fruto (MENDES e URBEN, 2015).
Controle:
Controle químico: Os benzimidazóis podem ser aplicados nos
pomares até três semanas antes da colheita; em pós-colheita, no Brasil, o uso
de fungicidas restringe-se aos benzimidazóis e ao imazalil (LARANJEIRA et al.,
2005). Três semanas antes da colheita pode-se efetuar pulverização das plantas
com Benzimidazóis (SANTOS e MENDES, 2015).
Controle físico: Armazenar as frutas em temperaturas abaixo de 5
°C, evitando o crescimento do patógeno que se desenvolve em temperaturas ideais
em torno de 24 ºC (UENO, 2011).
Controle
mecânico: fazer uma
desinfestação preventiva com produtos à base de cloro, amônio quaternário,
formaldeído ou álcool, evitando frutos infestados em veículos, equipamentos,
materiais de colheita, locais de armazenamento, de transporte e processamento
(ROSSETTI, 2001).
Controle
cultural: Manuseio
cuidadoso de frutos durante as operações de colheita, transporte, processamento
e armazenamento, visando evitar os ferimentos, que se constituem na principal
via de penetração do fungo em frutos; remoção imediata dos
frutos infectados, pelo uso de exaustores e de um bom sistema de limpeza local,
visando a diminuição de esporos do ar (LARANJEIRA et al., 2005);
Controle
biológico: as informações
sobre agentes biológicos que podem controlar o patógeno do bolor verde ainda
são muito restritas. Portanto, em um trabalho publicado por Kupper et al. (2013), mostra que é
viável o uso de Saccharomyces
cerevisiae para o controle de P. digitatum, porém a eficiência de antagonismo do
isolado depende da variedade cítrica que está sendo tratada.
Controle
genético: por ser uma
doença de pós-colheita causada por um patógeno relativamente fraco, que possui
ampla gama de hospedeiros, onde as medidas de controle citadas anteriormente
podem controlar a doença com facilidade, o uso de controle genético não é
utilizado. Mesmo porque este método é relativamente caro, e a doença não
necessita de tal tecnologia.
LITERATURA
CITADA:
AGROFIT
– Sistema de agrotóxicos fitossanitários. Ministério da Agricultura, Pecuária e
Abastecimento - Coordenação-Geral de Agrotóxicos e Afins/DFIA/DAS.
Disponível em: http://agrofit.agricultura.gov.br/agrofit_cons/principal_agrofit_cons. Acesso em: 6 de maio de
2015.
FARR, D.F., ROSSMAN, A.Y. Fungal Databases,
Systematic Mycology and Microbiology Laboratory, ARS, USDA. Retrieved June 26,
2015, from http://nt.ars-grin.gov/fungaldatabases/
FEITCHTENBERGER, E.; BASSANEZI, R. B.; SPÓSITO, M. B.,
BELASKE, J. Jr. Doenças
dos citros. In: KIMATI, H.; AMORIM, L.; REZENDE, J. A. M.; BERGAMIN FILHO, A.;
CAMARGO, L. E. A. Manual de Fitopatologia
vol.2: doenças das plantas cultivadas. 4ª
ed. São Paulo: Agronômica Ceres, 2005.
FORNER,
C.; BETTIOL, W.; NASCIMENTO, L.M.; TERAO, D. Controle
em pós-colheita de Penicillium
digitatum em laranja-pera com
microrganismos e tratamento térmico. Revista Brasileira de Fruticultura
v. 35 no. 1 Jaboticabal mar. 2013.
FRANCO,
D.A.S.; BETTIOL, W. Controle de bolor verde em pós-colheita de citros com
produtos alternativos. Empresa Brasileira de Pesquisa Agropecuária –
Embrapa Meio Ambiente no. 10 p. 1-4, fev. 2000.
INDEX FUNGORUM.
Banco de dados para táxons fúngicos. Disponível em:
Acesso em: 14 de Abril de 2015.
KUPPER, K.C.; CERVANTES, A.L.L.; KLEIN, M.N.; SILVA, A.C. Avaliação
de microrganismos antagônicos, Saccharomyces
cerevisiae e Bacillus subtilis para
o controle de Penicillium digitatum.
Revista Brasileira de Fruticultura, v. 35 no. 2 Jaboticabal Jun. 2013.
LARANJEIRA,
F.F.; AMORIM, L.; BERGAMIN FILHO, A.; AGUILAR-VILDOSO, C. I.; COLETTA FILHO, H.
D. Citros, cap. 18. Fungos procariotos e doenças abióticas. Cordeirópolis, SP:
FAPESP, 2004.
LARANJEIRA,
F.F.; AMORIM, L.; BERGAMIN FILHO, A.; AGUILAR-VILDOSO, C.I. Controle das
doenças causadas por fungos e bactérias em citros. In: ZAMBOLIM, L.; VALE, F.
X. R.; MONTEIRO, A. J. A.; COSTA, H. Controle de doenças de plantas fruteiras. Vol 1, Minas Gerais: UFV, 2002.
MENDES,
M.A.S.; URBEN, A. F.; Fungos relatados em plantas no Brasil, Laboratório de
Quarentena Vegetal. Brasília, DF: Embrapa Recursos Genéticos e Biotecnologia -
CENARGEN. Disponível em: .
Acesso em: 6 de maio de 2015.
ONIONS, A. H. S.
Penicillium digitatum. C.M.I.
Description Of Pathogenic Fungi and Bacteria no. 96. Commonwealth Agricultural
Bureaux, June 1966.
PATRO, Raquel. Laranja – Citrus
sinensis. Disponível em: http://www.jardineiro.net/plantas/laranja-citrus-sinensis.html.
Acesso em: 21 de abril de 2015.
ROSSETTI, V. V. Manual
ilustrado de doenças dos citros. Piracicaba: Fealq/Fundecitrus, 2001.
SAMSON, R.A.; HOUBRAKEN, J.; THRANE, U.; FRISVAD, J.C.; ANDERSEN, B. Food
and indoor fungi. CBS – KNAW –
Fungi Biodiversity Centre Utrecht, The Netherlands, 2010.
UENO, B. Produção orgânica
de citros no rio grande do sul. Embrapa Clima Temperado, dezembro 2011.
Disponível em: http://www.cpact.embrapa.br/publicacoes/sistemas/sistemas-20/cap10_doencas_dos_citros.htm. Acesso em: 12 de maio
de 2015.